
Podkmen Crinozoa Matsumoto, 1929
(řec. krinos - lilie, zóon - zvíře)
Zahrnuje tyto třídy: Crinoidea a Edrioasteroidea
Crinoidea Miller, 1821 - lilijice/
výskyt spodní ordovik - recent
ROZCESTNÍK
Crinoidea
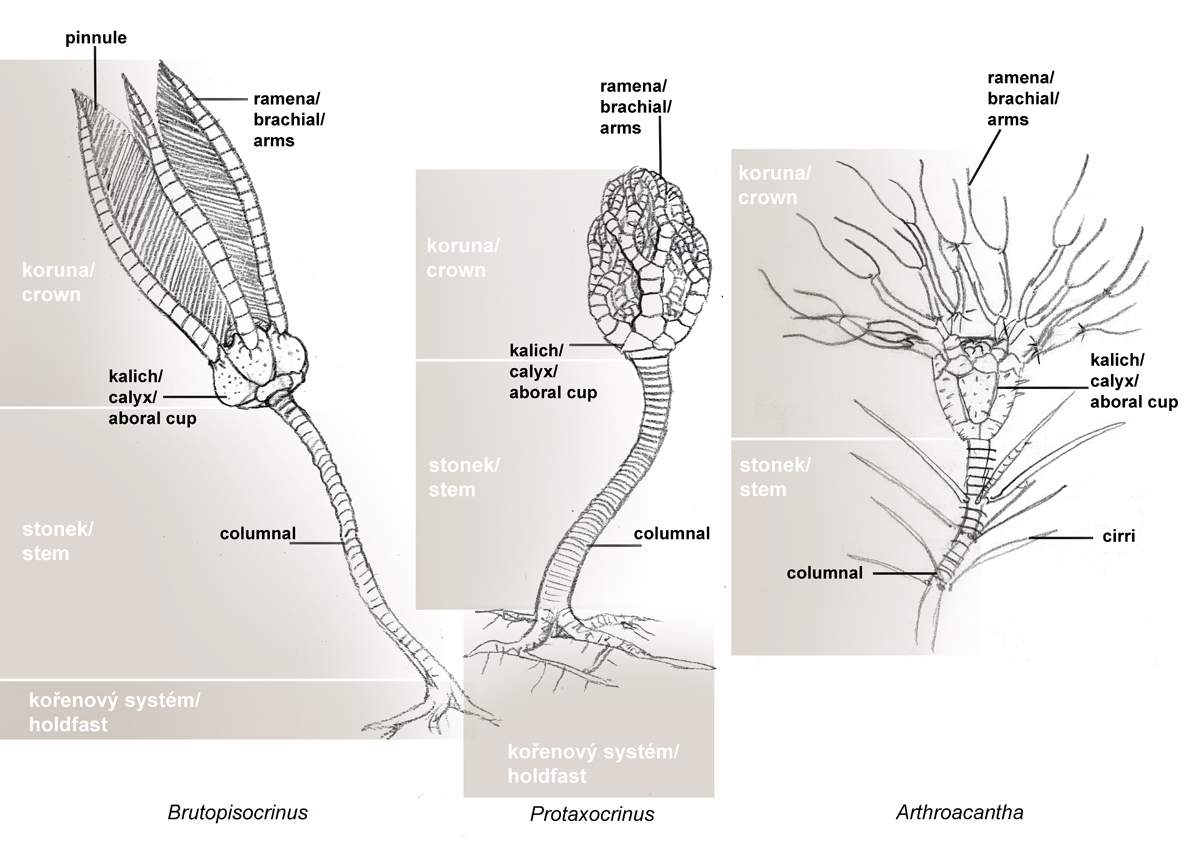
Obr. 1 - základní morfologie vyhynulých přisedlých lilijic.
Třída Crinoidea je velká, tvarově bohatá větev ostnokožců, která se vyskytuje od svrchního ordoviku do současnosti. V prvohorách tato třída zahrnovala tři gigantické podtřídy: Camerata, Disparida (dříve Inadunata) a Flexibilia. Někteří autoři nesouhlasí s tímto dělením a všechny zahrnují do gigantické vymřelé podtřídy Cladida. Mnozí autoři tuto třídu dělí na podtřídy čtyři s tím, že podtřídu Cladida zařadili ke třem stávajícím - Camerata, Disparida, Cladida a Flexibilia. Ať už tyto čtyři, tři, či tato jedna podtřída měly stovky rodů s více než 2 500 druhy - nové se stále nalézají. Vrcholu rozvoje dosáhli během karbonu, ale již na počátku permu můžeme zaznamenat nápadný ústup jejich druhového bohatství i početního zastoupení a koncem permu zástupci těchto archaických podtříd mizí ze světových moří. Vlastně až na dvě výjimky. První je druhohorní rod Encrinus, který někteří badatelé zařazují ještě k prvohorním inadunátním, respektive disparidním lilijicím. Druhou výjimkou je recentní rod, dříve považovaný za vyhynulý Guillecrinus. V roce 1982 bylo u ostrova Reunion v Indickém oceánu vyloveno pět exemplářů druhu Guillecrinus reunionensis. Jejich objev byl vědeckou senzací. Téměř všichni paleozoičtí krionidi totiž koncem prvohor vymřeli a krionidi dnešních moří (cca. 700 zatím známých druhů) patří vesměs do "moderní" čtvrté podtřídy Articulata, která se objevuje až začátkem druhohor, v triasu a na tři předešlé podtřídy bezprostředně navazuje.
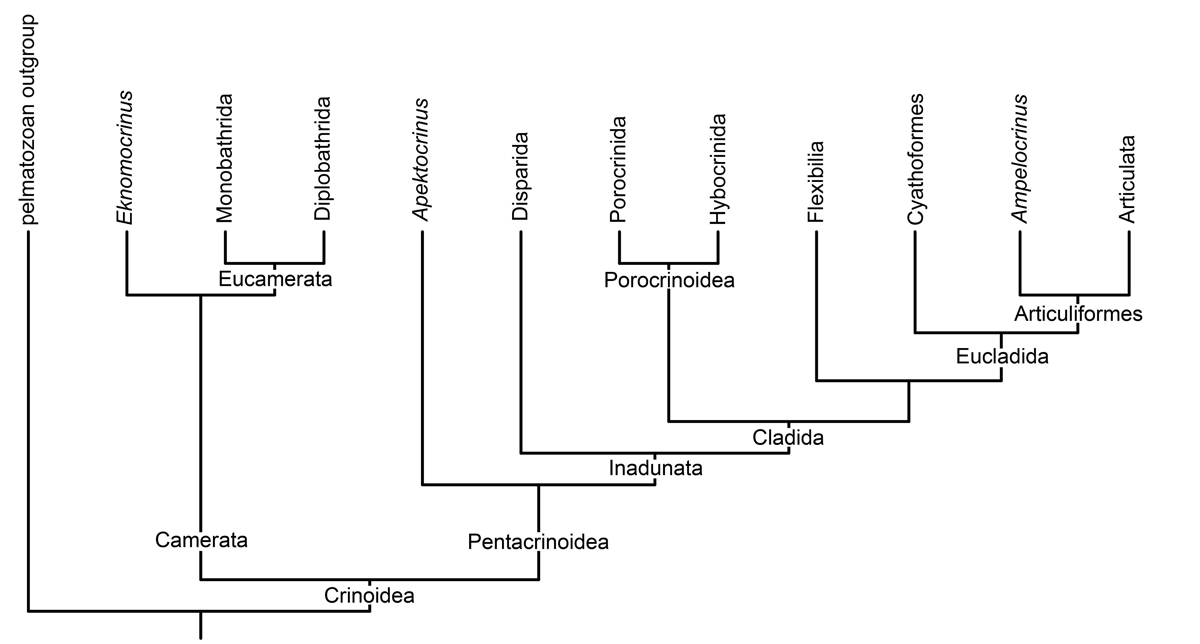
Obr. 2 - schématická tabulka jednoho z mnoha systémů dělení krinoidů.
Crinoidea je možné označit jako vývojově i stavbou těla specializované ostnokožce s většinou pravidelně utvářenou tékou, rozdělenou na kalich, článkovaná ramena, článkovaný stonek s upevňovacím, kotevním zařízením, v několika případech orgánem plovoucím. Kalich a ramena tvoří dohromady tzv. korunu, někdy tak podobnou jakémusi fantastickému květu, že by laik těžko poznal, že jde o živočicha. Máme malé podezření, že se švýcarský surrealistický fantasy umělec H. R. Giger nechal inspirovat při tvorbě vetřelce, konkrétně jeho vývojovou fází tzv. facehugger, právě kalichem lilijice s chapadly. Není nic neobvyklého, ani ojedinělého, kdy se nějaký scifi tvůrce nechal inspirovat vyhynulou formou. Dýchací soustava a smysly zcela chybí.
KALICH A RAMENA
Na vrcholu stonku (který může být velmi dlouhý, až 25 metrů) je kalich (theca, někdy také calyx), který obsahuje měkké části těla. Z horního okraje kalicha vybíhá pět a více pohyblivých, velmi často dále větvených ramen (brachia). Ramena rozeznáváme jednoduchá - uniseriální, kdy leží jednotlivé klouby za sebou a zdvojená/dvouřadá - biseriální, kde mají jednotlivé klouby tvar klínů a prolínají se střídavě doprava a doleva . Přechod od uniseriálních k biseriálním na obrázku 7a (podle Bathera a Liljevalla, 1893). V prostorách mezi bázemi ramen vybíhá okraj kalicha směrem do středu v tzv. střechu kalicha (tegmen), uprostřed níž je ústní otvor a extracentricky (v interradiu) otvor anální. Trávící soustava je ve tvaru U. Stěny kalicha jsou tvořeny pětičetnými prstenci destiček, které spolu vzájemně alternují. Nejvýše položený zahrnuje pět radialií, k nimž se kloubí článkovaná ramena. Některá ramena jsou opatřeny pinulemi (pinnule = ambulakrální nožky), tedy jakýmisi lepivými, pohyblivými chapadélky, které slouží ke snadnějšímu úchytu potravy a následného transportu k ústnímu otvoru. Ten je často ukryt pod destičkami tegmenu. U některých druhů je, jak již bylo zmíněno výše, ústní otvor ve středu tegmenu, anál je středově vychýlen. U některých druhů se zdá, že je to naopak, přičemž anální otvor hypertrofoval v tzv. anální tubus. Je to ale spíše tím, že vnik análního tubusu ústa pouze vychýlil ze svého středu. Pod prstencem radialií leží bazální prstenec, tvořený pěti interradiálně umístěnými bazálními destičkami (basalia). Rozeznáváme dva základní typy kalichu - monocyklický a dicyklický (viz. obr. 3).
Vzájemný poměr kalicha a ramen je velmi rozmanitý. V kalichu jsou uloženy základní tělesné orgány (ústa, řitní otvor nebo trubice). Ramena byla často bohatě větvená, volně pohyblivá (alespoň jejich vrcholové části). Jejich úkolem bylo zachycovat mikroskopickou potravu a dopravovat ji za pomocí pinulí k ústnímu otvoru.
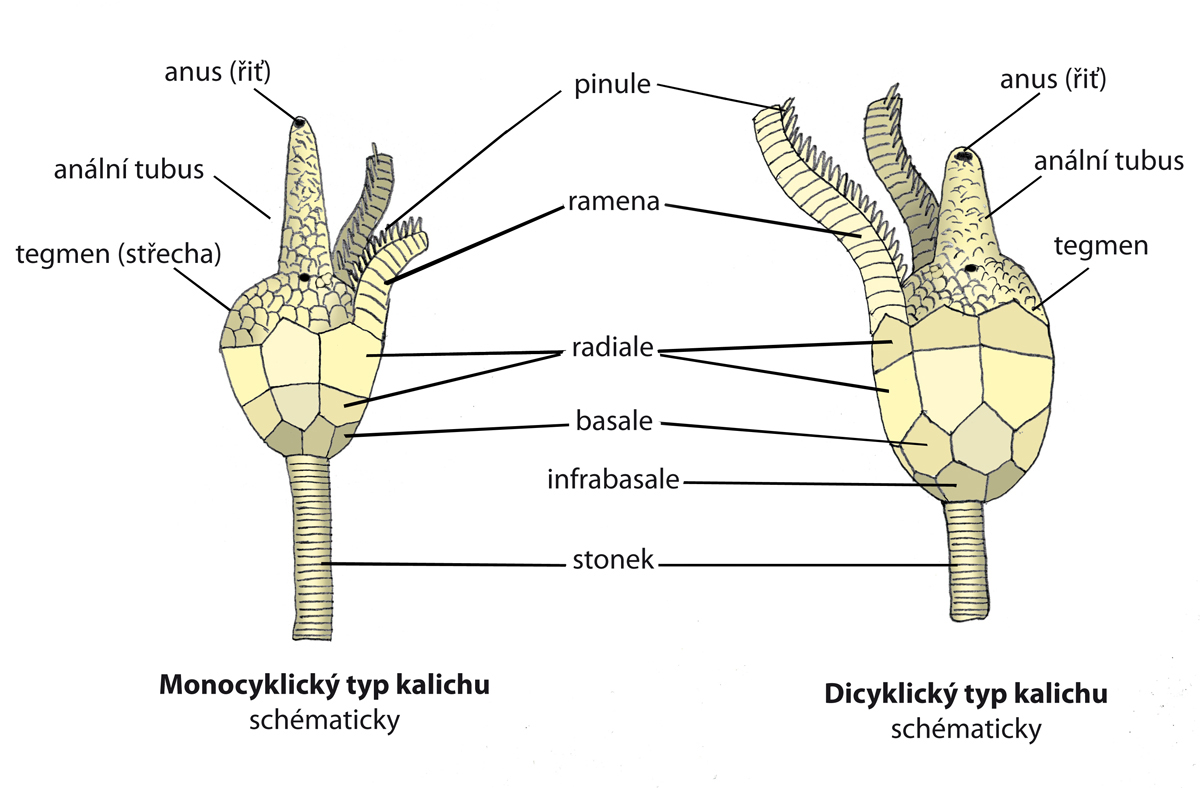
Obr. 3 - základní schématická morfologie kalichu monocyklické a dicyklické lilijice. Ne všechny lilijice byly opatřeny např. análním tubusem. U některých typů ležel anus vedle ústního otvoru, který je zde zobrazen u ústí tubusu tmavou skvrnou. Jak je zřejmé, monocyklickým typům kalichu schází infrabazální tékální destičky a na stonek navazují přímo bazálními destičkami.

Obr. 4 - morfologie kalichu (téky) kamerátní lilijice druhu Stereocrinus triangulatus Barris (nahoře) a druhu Dolatocrinus icosidactylus Wachsmut et. Springer (dole) na vyobrazení podle Ch. Wachsmuta a F. Springera - The North American Crinoidea Camerata, Cambridge, 1897.
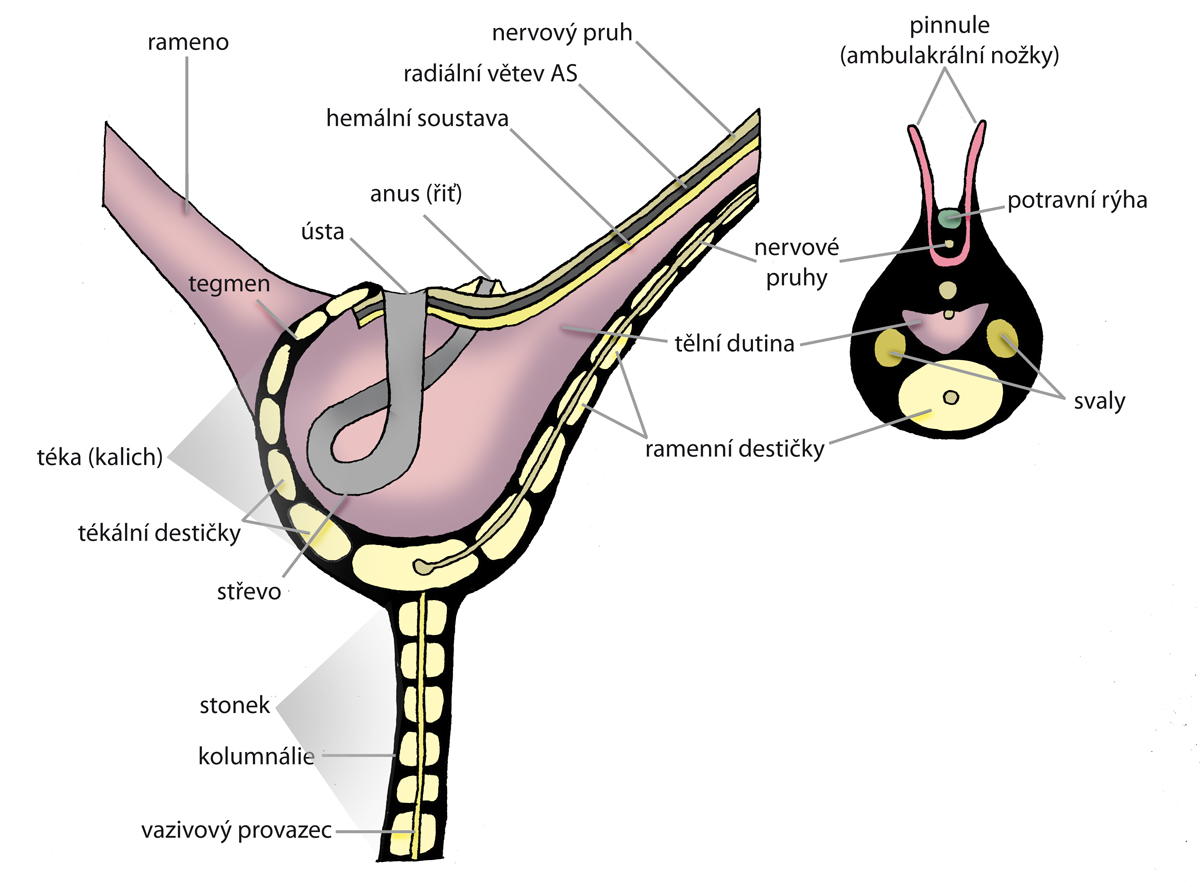
Obr. 5 - anatomie vnitřní stavby těla lilijic. Vlevo kalich přisedlé lilijice, vpravo průřez ramenem. Překresleno podle M. Mergla - Západočeská univerzita v Plzni, Fakulta pedagogická, Centrum biologie, geověd a envigogiky.

Obr. 6 - základní typy kalichů lilijic podle Ubaghse, 1978.

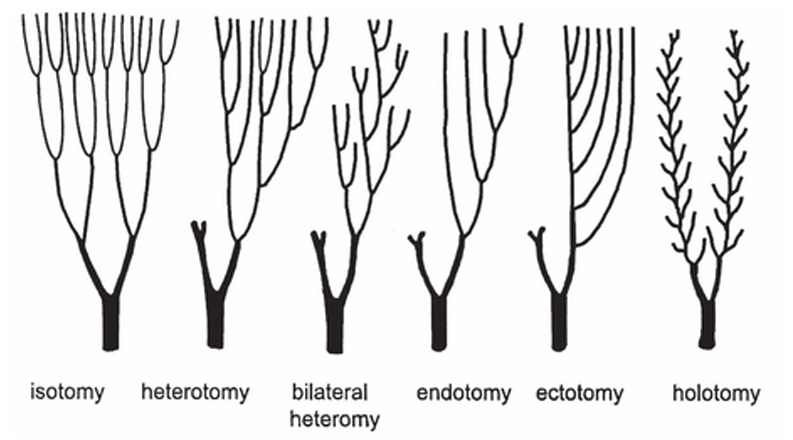
Obr. 7a - přechod od uniseriálních ramen k biseriálním
7b - základní typy větvení ramen lilijic podle Ubaghse, 1978.
STONEK
Stonek (vyvinutý u většiny prvohorních lilijic) je více či méně ohebná část téky složená z nízkých, okrouhlých, eliptických, čtverhranných, pětiúhelníkovitých nebo hvězdicovitých článků (kolumnálie), uspořádaných v jedné řadě nad sebou. Uprostřed každého článku - kolumnálie, je nejčastěji kruhový nebo hvězdicovitý, méně častěji podkovovitý otvor, či jinak tvarovaný otvor pro svalový a nervový provazec (lumen). Po jeho krajích je často vyvýšená část (perilumen). Po krajích kolumnálií je často jakýsi ozubený zámek (crenulae = crenularium). Ten může mít mnoho podob a forem. U některých typů však tento zámek schází nebo je zanedbatelný. Občas se crenulae paprsčitě sbíhá až ke středovému otvoru, tedy lumenu. U některých typů však přechází v hladkou nebo lehce pórovitou plochu (areola). Obrázky 8. - 11.
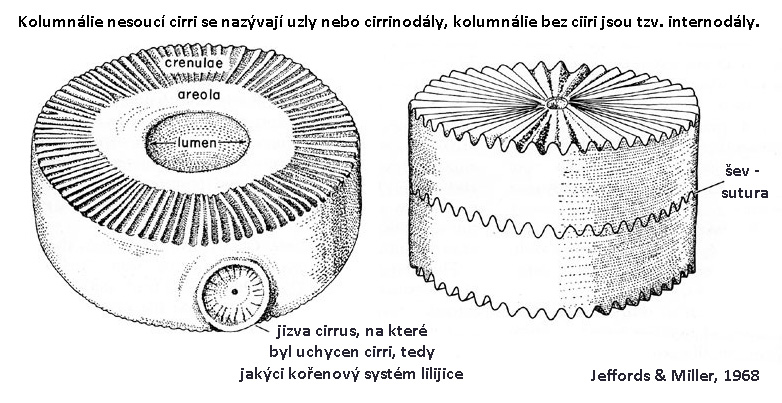
Obr. 8 - stavba cirkulární kolumnálie podle Jeffordse a Millera, 1968.

Obr. 9 - stavba pentagonální kolumnálie podle Carpentera, 1884.

Obr. 10

Obr. 11 - schématická stavba kolumnálie.
Tři základní typy švů na stoncích lilijic Jednotlivých typů stonků je nepřeberná řada, která se odvíjí od samotných kolumnálií, které ne vždy byly kruhového, tedy nejběžnějšího řezu. Často jsou "půdorysu" pentagonálního, čtverhranného, či dokonce výrazně asymetrického. Tady jde však ale spíše o tři základní morfologické švy (sutury), tedy o ukotvení, či jakýsi zámek, kterým byly jednotlivé kolumnálie k sobě přichyceny. Překresleno podle Donovana, 1989.
Tvarová diverzita kolumnálií lilijic je neuvěřitelně různorodá a není snadné se v ní orientovat. Často se tvar, velikost a morfologie kolumnálií proměňuje i v rámci jednoho jediného stonku, tedy že jiné kolumnálie jsou u ukotvovacího aparátu, které se mění směrem k théce krinoida. Kolumnálie se proměňovaly také v rámci vývojové ontogeneze lilijice (obr. 12). Tato různorodost je krásně vidět na koláži níže (obr. 13), na které je pouhý zlomek, kapka v moři, z mnoha typů kolumnálií různých druhů krinoidů.
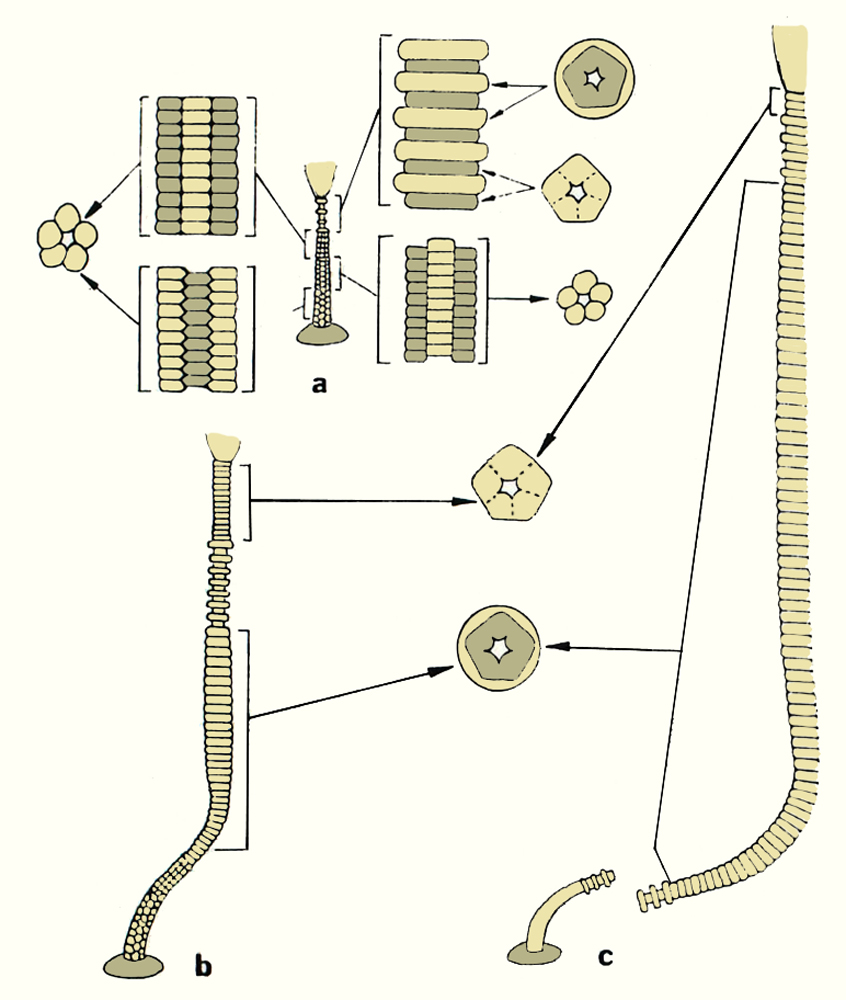
Vývojová ontogeneze stonku ordovické disparidní lilijice druhu Cincinnaticrinus varibrachialus Warn and Strimple 1977 † (Kanada, USA) od juvenilního stadia (a), až po stadium dospělosti (c).

Obr. 13 - tvarová diverzita několika málo z mnohem většího množství kolumnálií napříč druhy i časem - od ordoviku až po recent.
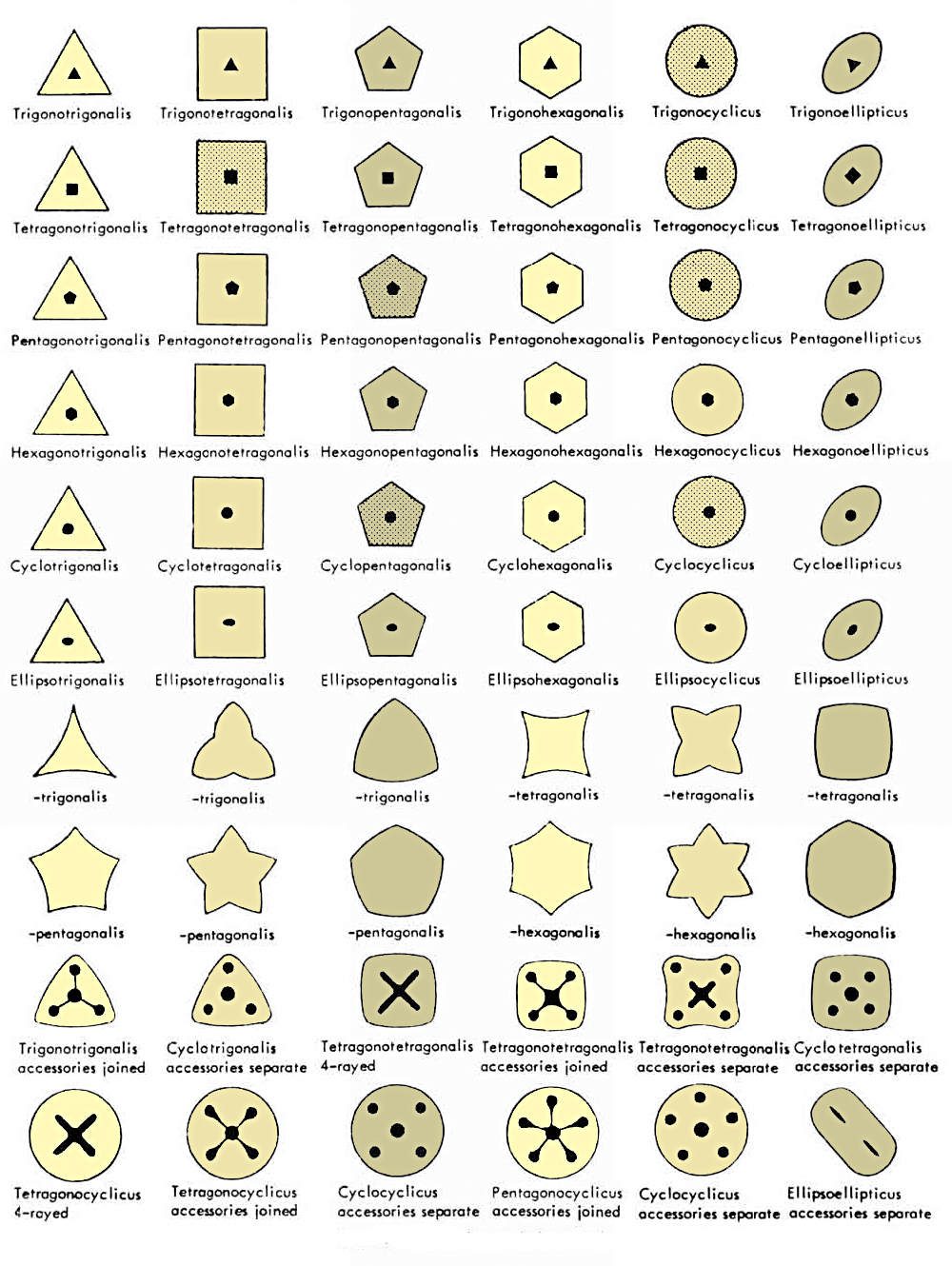
Obr. 14 - umělá klasifikace krinoidních sloupců a pro ně navržená nomenklatura - YELTYSHEVA (1955, 1956, 1959, 1964). Z části ji používají i další autoři ve svých systematických popisech např. (YELTYSHEVA, 1955, 1960, 1964; YELTYSHEVA & DUBATOLOVA, 1960 (1961); YELTYSHEVA & STUKALINA, 1963; SIZOVA, 1960; AVROV & STUKALINA, 1964; DUBATOLOVA, 1964, 1967; DUBATOLOVA & SHAO. 1959, DUBATOLOVA & YELTYSHEVA, 1961a, b; STUKALINA, 1960, 1961, 1965a, 1966). Sloupce s jednoduchými typy axiálních kanálů jsou v horní části diagramu, změny v příčném tvaru a komplexní axiální kanály ve spodní části diagramu.
Samotný stonek je složen z mnoha jednotlivých, do sebe zaklesnutých a ve většině případů pohyblivých kolumnálií = internodály, jejichž počet převažuje nad nodály nebo také cirrinodály (jakési uzliny) = plurikolumnálie, ze kterých často rostly výběžky, které nazýváme cirri, tedy jakýsi pomocný ukotvovací aparát. Ne vždy jsou tyto plurikolumnálie ale nositeli cirri (obr. 15). U dnešních stopkatých lilijic jsou stonky celkem krátké, okolo 20 cm, ale u vyhynulých druhů se jeho délka měnila od několika málo cm až do téměř 25 m! Totéž platí i pro tloušťku, tedy průměr (u mikrolilijic je to pod 1 mm, u gigantických druhů je to v průměru 15 - 20 cm).
Jistý, ale poněkud jiný extrém v tomto smyslu představují lilijice, u nichž kalich se stonkem hypertrofuje v neforemnou hmotu, jež spodinou (velmi silným kořenovým systémem) přisedá ke skalnímu dnu. Tyto lilijice se vyznačují velmi krátkými stonky, ale extrémně silnými. Často jsou gigantických rozměrů a jejich stonky připomínají spíše sloní choboty (tak je také dělníci pracující v Koněprusích s oblibou pojmenovávali). Za všechny snad jen rody Pernerocrinus, pojmenovaný dle titána české paleontologie Pernerovi nebo Perunocrinus pojmenovaný dle titána slovanské mytologie boha hromu a blesku Peruna. Druh Torrocrinus grandis Prokop et Petr, 1991 † z koněpruských vápenců pražského souvrství si své jméno opravdu zaslouží. Tento druh byl dokonce tak mohutný, že ač jeho stonek byl průměrně vysoký pouhých 10 cm, tak jeho brachiální průměr byl opravdu grandiózní a to pravděpodobně až ale dva a půl metu!. Samozřejmě se takový stonek v Koněprusích nenašel kompletní, ale ojedinělé nálezy tohoto rodu a druhu učiněné Rudolfem J. Prokopem, této předpokládané velikosti velmi, velmi nasvědčují. A to také nemuseli nalézt největší gerontické jedince.
I když jsou stonkoví krinoidi dobře známí z moderních oceánů, post-paleozoická podtřída Articulata představuje pouze nepatrné zastoupení, které přežilo zánik mnoha dalších cladů, tvořících taxonomický "strom", jež ale nedokázaly přežít masové vymírání na konci Permu. Třebaže mnoho formálních a ekologických aspektů
existujících stonkových krinoidů může pomoci vysvětlit funkční morfologii a mezerovou diferenciaci paleozoických taxonů, mezi živými členy skupiny existuje jen velmi málo blízkých analogií. Většina paleozoických krinoidů má totiž morfologické rysy bez svého moderního protějšku. To platí zejména o stonku/sloupci i struktuře připevnění (upevňovacím aparátu), který měl daleko větší variabilitu tvaru, než bylo zjištěno z post-paleozoika. Primitivní ležící krinoidi mají obvykle homeomorfní proxistelu, která může být také rysem vzpřímených lilijic, s pouze slabě reofilním habitem. Primitivní sloupce/stonky s širokými kruhovými axiálními kanály mohou být "drženy" po předcích zděděnými tzv. Hohlwurzeln ("dutými kořeny"). Tento dutý prostor mohl být vyplněn perihaemální kapalinou, ale také mohl obsahovat další, výživnější tkáně přitažlivé pro predátory nebo mrchožrouty.

Obr. 15 - A) Austinocrinus mexicanus - křída, Mexiko, B) Apiastrum candidum - devon, Tennessee, C) Graphosterigma synthetes - Misssissippian, Kentucky, D) Cylindrocauliscus fiski - Misssissippian, Texas, E) Cyclocaudex plenus - Pennsyvanian, Texas.
KOŘENOVÝ SYSTÉM - UKOTVOVACÍ APARÁT
Velmi zajímavý je i ukotvovací aparát - jakýsi "kořenový systém" nazývaný cirri. Zatímco pralilijice - eokrinoidi byli často (až na výjimky) ukotveni k podloží jakýmsi biologickým lepidlem (biogluing), tak lilijice si vyvinuli poměrně sofistikovaný a velmi variabilně rozvinutý systém ukotvení k podloží. Existuje několik videí, kdy se soudobé lilijice volně pohybují za pomocí ramen, ač velmi nemotorně, například zde a zde. Tyto recentní lilijice mají často rozpětí ramen kolem 7 metrů. Videa, kde neexistuje měřítko jsou tak velmi zkreslující, protože vypadají, že jsou ve skutečnosti drobnější. Tuto schopnost volného pohybu za pomocí ramen, se ale u archaických druhů nepodařilo ověřit a prokázat, není však vyloučena. U soudobých, volně plujících lilijic bylo zjištěno, že v mladém věku ještě stonkem disponují, aby jej posléze v dospělosti oddělily a tímto způsobem se pak volně pohybovaly.
Co se týče vymřelých fosilních druhů, tak u mnohých krinoidů tento systém opravdu připomíná mělký kořenový systém recentních nahosemenných rostlin. U jiných krinoidů není tento systém jenom výsadou stonkové podsady, ale kořenová ukotvení vyrůstala mnohdy až do poloviny stonku (často výše) samotné lilijice (viz. Devonští crinoidní ostnokožci z Barrandienu - Camptocrinus, Platycrinites, Simakocrinus aj.). U těchto rodů jde o tzv. cirrinodály nebo prostě jen nodály, tedy nosiče kořenů. Ty ostatní články jsou tzv. internodály. Logicky jde o krinoidy, kteří se denně potýkali s příbojovými nesnázemi a museli být pevně ukotveni buďto k podloží, to je v případě, kdy žili ve štěrbinách útesů, ke kterým mohli být přichyceni po téměř celou délku stonku k matrici/podloží. V druhém případě byli pomocí cirriri ukotveni jeden ke druhému a snázeji tak vytvářeli odolné kolonie.

Obr 16 - karbonský druh Camptocrinus multicirrus Springer, 1926 ze stupně Mississippian, který své druhové jméno multicirrus, tedy mnoho cirrusový, nemá rozhodně náhodou. Tento druh je pro cirrusové "přívěsky" zcela typickým zástupcem. Zde, na naší idealizované rekonstrukci je schématicky znázorněn v předpokládaném životním prostředí, přičemž jsme chtěli pomocí této kresby zároveň vysvětlit, k čemu že to ty cirri, tato chapadélka, vlastně mohly sloužit. Za prvé zřejmě k nějakému úchytu k podloží. V tomto případě se nabízí nějaká štěrbina ve skále, kde bylo příbojové moře, aby příboj poměrně subtilního živočicha zcela neroztrhal a nerozbil. Za druhé mohli být cirri jednoho živočicha propleteny s cirri jiného, aby vytvořili kolonii, jakousi pospolitos, která je obecně více odolná než solitérní jedinec. S tím samozřejmě souvisí za třetí, tedy nějaká komunikace mezi samotnými jedinci. Je to hodně scifi, jelikož víme, že lilijice se řadí k nejprimitivnějším ostnokožcům, kteří jsou bytostní dravci a jež nemají smysly, ale co my víme jakou měli v minulosti loveckou strategii, kdy jich žilo takové množství. Mohli ve skutečnosti disponovat třeba i nízkým vědomím. Lidé měli vždy tendenci v tomto směru živé bytosti podceňovat. Dnes se naopak spekuluje, že i rostliny spolu komunikují na úrovni jistého vědomí. A k čemu jim tedy sloužili cirri? Zcela jistě měly svůj význam, který se v moderní době stal pochybným zde. Na videu je recentní lilijice, která se pohybuje pomocí ramen koruny po dně moře. Za korunou vláčí poměrně dlouhý stonek/sloupec, který je opatřen cirri. Ty jí však evidentně k pohybu neslouží, naopak to vypadá, že celý stonek i s přívěsky je jí překážkou, jakousi vězeňskou koulí minulosti.
Zcela mimořádnou roli, ač to byla zřejmě odchylka, která se již nezopakovala, měli kamerátní lilijice čeledi Scyphocrinitidae Jaekel, 1918 †. Tyto lilijice dosáhli na přelomu svrchního siluru a spodního devonu takového úspěchu, že se staly v tehdejších mořích jednou z nejúspěšnějších skupin živočichů. Z našich končin je znám zvláště druh Scyphocrinites elegans Zenker, 1833, významný horninotvorný živočich např. z chráněné lokality Budňanská skála u Karlštejna. Příčinou jejich celoglobálního "masového" výskytu byla evoluční odchylka od ostatních krinoidů. Tím byl svého času měkký, vakovitý, několikakomorový orgán zvaný lobolit (obr. 18), neboli plovák, kterým byl ukončen jejich stonek (místo "kořenového" aparátu). Tento plovák umožňoval scyfokrinitům, po naplnění plynem, který tyto lilijice produkovaly, se volně vznášet a unášet mořskými proudy. Po naplnění vodou naopak posloužil jako kotva, kdy mohly tito krinoidi pevně usednout k podloží. Dnes již defakto zaniklá a vytěžená lokalita v Prokopském údolí se stále nazývá, dle vysokému počtu historických nálezů těchto plováků, jako "Lobolitová stáň". Nejvýznamnější lokalita nálezů Skyfokrinitů však nadále náleží Budňanské skále. Velmi dobré naleziště je i v Žákovu lomu v Malé Chuchli u Prahy.
Poslední dobou se vyrojili nálezy, patrně kamerátních, nicméně blíže neurčených lilijic z ordoviku Maroka, které jsou sloupcem napojeny přímo na lobolit. Pakliže se nejedná o fake, což může z 99% být* a vědecká obec se k tomu prozatím také nevyjádřila, znamenalo by to, že lobolity lilijice používaly již od středního ordoviku (obr. 17).
Co se týče jakési kotvy, tak tou disponoval v pravém slova smyslu rod Ancyrocrinus Hall 1862 (obr. 19 - A), kterou byl schopen "zakotvit" nejenom o překážky ve formě kamenů, živočichů, ale i například na dně písčitého moře. Tento rod lilijice se však u nás doposud nenašel. Zcela specifickou roli zde hrají pak určité rody mezzozoických lilijic, které "symbioticky" přisedly na kus spadlého kusu stromu, jež plul po hladině moře a tímto způsobem se nechaly transportovat z jedné strany oceánů na druhou. Byly nalezeny výjimečné fosílie, kdy na mrtvém kmenu stromu byly zachyceny celé takovéto kolonie lilijic, čítající mnoho jedinců (viz text níže).
* z podstaty velkého množství padělků z Maroka jsme k podobným nálezům značně nedůvěřiví

Obr. 17 - Marocká ordovická lilijice napojená na lobolit - fake/reál?
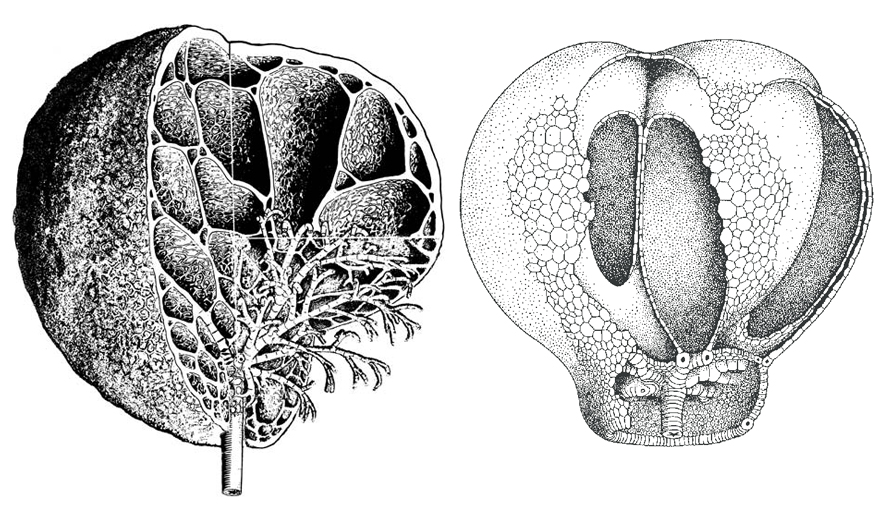
Obr. 18 - dva typy cirrusových lobolitů v pravděpodobné životní pozici podle Hesse, 1972 a 1979. Vlevo rod Scyphocrinites, vpravo rod Camarocrinus.
Obr. 19 - některé z typů kořenových systémů (cirri) lilijic.
A) devonská lilijice Ancyrocrinus bulbosus Hall, 1862 - kotva, B) silurská lilijice Eucalyptocrinites ovalis (Hall), C) devonská lilijice Ammonicrinus doliiformis Wolburg, 1938, D) jurská lilijice Liliocrinus munsterianus (d'Orbigny), E) ordovická lilijice Caleidocrinus (Huxleyocrinus) turgidulus Ramsbottom 1961, F) silurodevonská lilijice zavěšená na lobolitu Scyphocrinites elegans Zenker, 1833.
Díky velmi příznivým nálezům byly některé druhy krinoidů nalezeny přichyceny na plovoucím dříví. Vyplývá z toho, že lobolit tedy nebyl jediným způsobem transportu lilijic (obr. 18 a 19). Nejčastější výskyt tohoto jevu pochází ze svrchního devonu, tedy v době, kdy se u lilijic už lobolity nevyskytovaly, nejspíše byly slepou větví. Klug a kol. (2003) popsali velké množství malých, diskoidních, krustózních "kořenových úchopů" - holdfasts blíže neurčených krinoidů na kusech dřeva rodu Archaeopteris Dawson, 1871 v tmavě zbarvených "bahnitých" břidličných sedimentech východního Antiatlasu (severozápadní Maroko) - st. famen. Jde například, dle C. Kluga o druh lilijice druhu Mrakibocrinus bockwinkeli Webster, Becker & Maples, 2005 †, kdy naplavené dřevo leží přímo na rozptýlených krinoidních kůstkách výše zmíněného krinoida. Toto opakované spojení krinoidních kolonií s dřevěnými špalky lze interpretovat jako další příklad pseudoplanktonického způsobu života devonských krinoidů. Podobné výskyty jsou známy od svrchního stupně frasn po spodní famenské vrstvy v Severní Americe, kde byly lilijice, přiřazené k rodu Melocrinites Goldfuss, 1831 †, také připojeny radikály ke dřevěným špalkům (Wickwire, 1936; Welllls, 1942, 1947; McIntosh, 1978). Daleko přesvědčivější nálezy jsou známy z mezzozoika (druhohor - střední trias až spodní jura). Konkrétně jde o druhy řádu Encrinida Matsumoto, 1929 †(Traumatocrinus caudex Dittmar, 1866 †, Traumatocrinus hsui (Mu, 1950 †), případně Vostocavocrinus boreus †) a druhy řádu Isocrinida Sieverts-Doreck 1952, podřádu Pentacrinitina, konkrétně druhy Pentacrinites fossilis Blumenbach, 1804 †, Pentacrinites dichotomus (McCoy, 1848 †) a Seirocrinus subangularis (Miller, 1821 †) ze spodní jury. Druh Seirocrinus klikushini Simms, 1989 † a blíže nepopsaný nález pentacrinidů ze svrchního triasu (severní Sibiř).


Obr. 20 - transport kolonie krinoidů druhu Traumatocrinus hsui (Mu, 1950 †) na kusu dřeva. Koruny jsou podobné velikosti, ale stonky se liší délkou tak, aby dosáhly různých hloubek, délka naplaveného dříví 3, 3 m, formace Xiaowa, Ladinian-Carnian boundary, Guanling, Čína (podle Hagdorn & others, 2004). Obr. 21 - transport kolonie krinoidů druhu Pentacrinites fossilis Blumenbach, 1804 † a jejich umístění na kmenu stromu. Čím blíže k hladině, tím častěji jsou přítomny juvenilní formy krinoida (Simms, 1999).
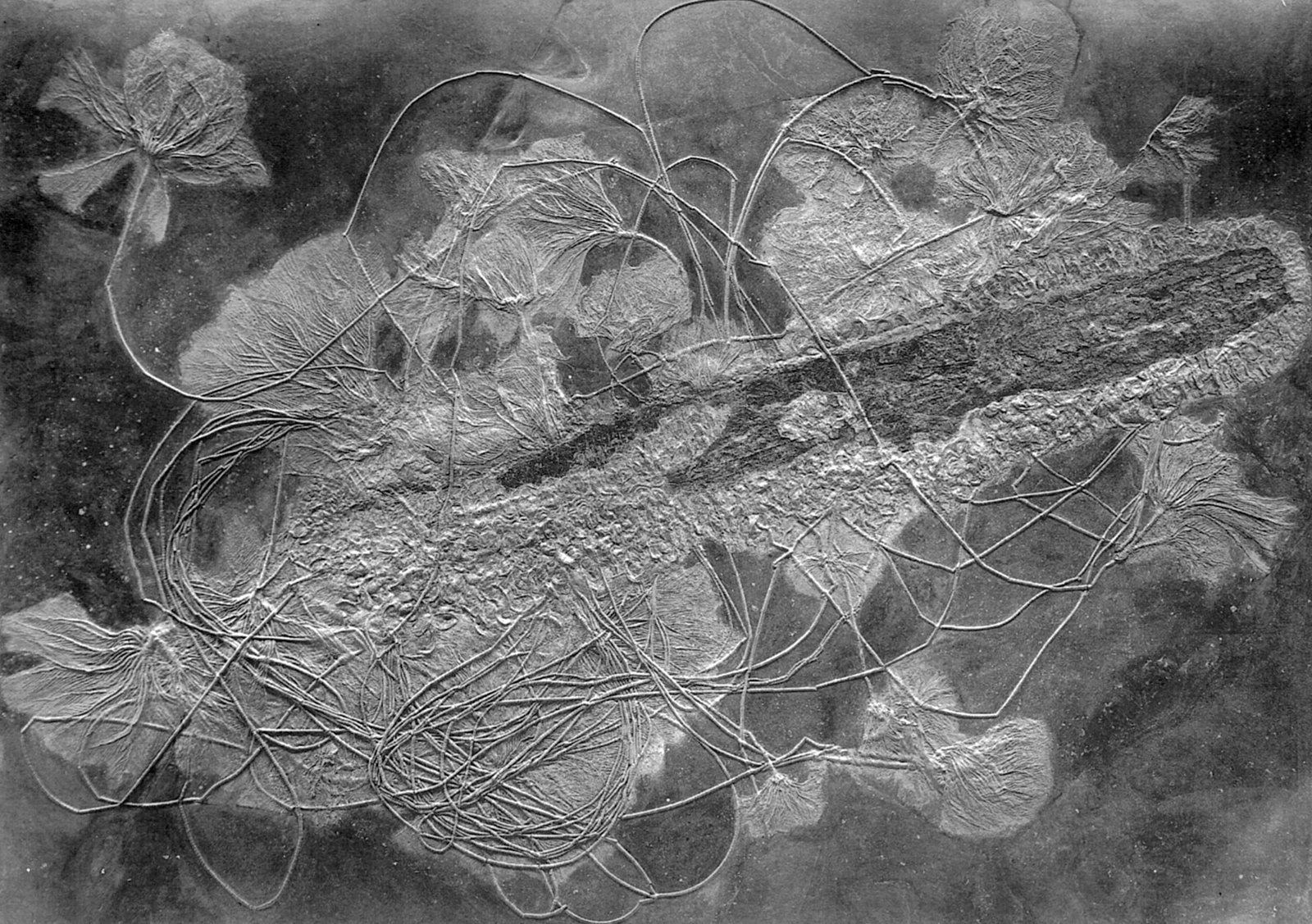
Jak takový unikátní nález, tedy kolonie lilijic, které jsou přichycené na plovoucím transportním kusu dřeva v reálu vypadá si můžete prohlédnout na fotografii níže. Jedná se o lilijice jednoho druhu Seirocrinus subangularis (Miller, 1821 †) ze spodní jury, které jsou ukotveny ke kusu kmenu stromu nejistého zařazení, jež je zároveň pokryto mlži rodu Pseudomytiloides Koschelkina 1963 †, stupeň Toarcian (spodní jura), Posidonia Shale, Holzmaden, Německo (Hauff Museum, Holzmaden).
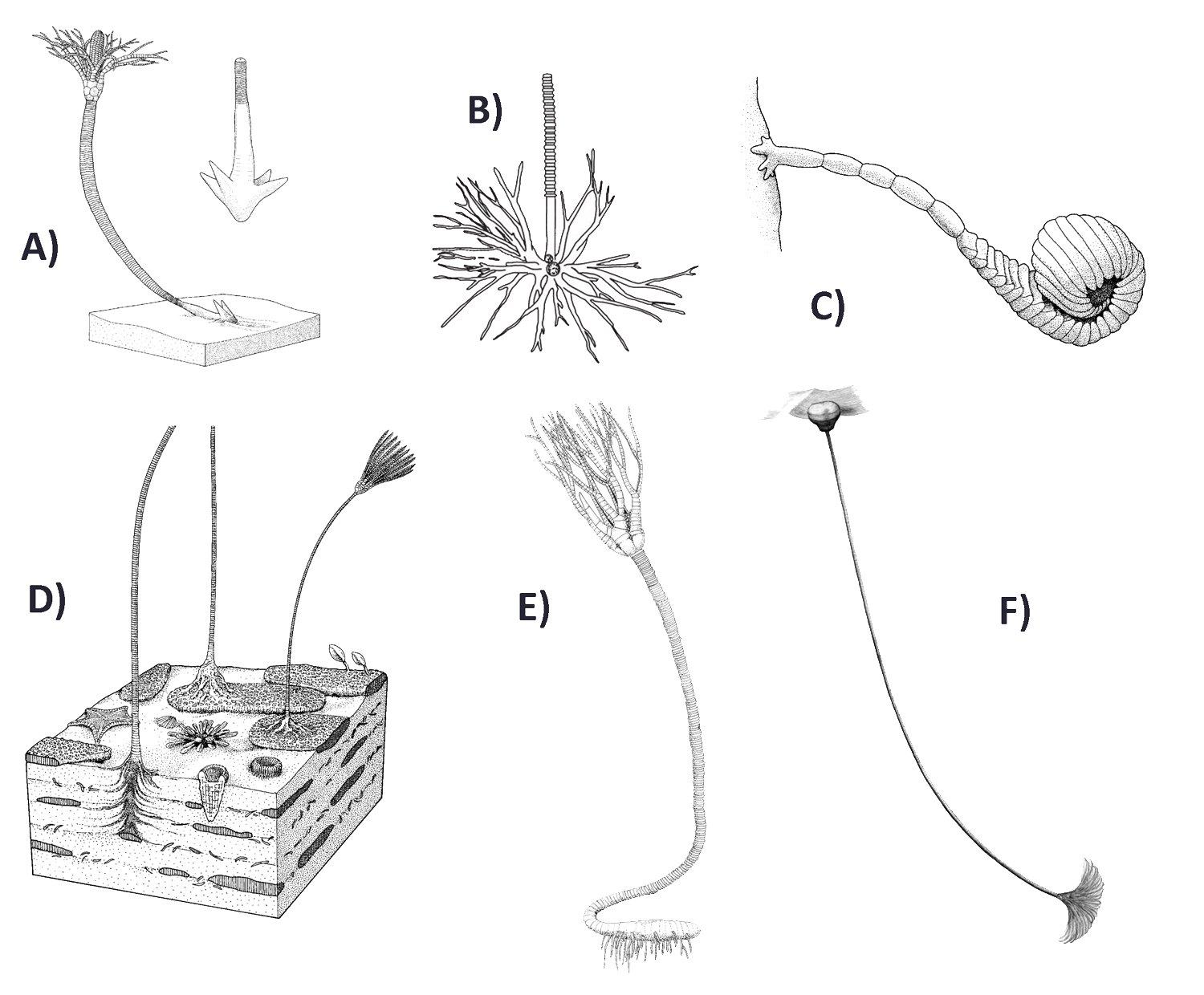
Čtyři základní podtřídy krinoidů
Významné jsou tyto rody - Scyphocrinites ZENKER, 1833, Platycrinites Miller 1821, Agariocrinus (u nás prozatím zástupce tohoto rodu nenalezen).

Obr. 23 - ukázka stavby těla kamerátní lilijice dle G. Ubaghse
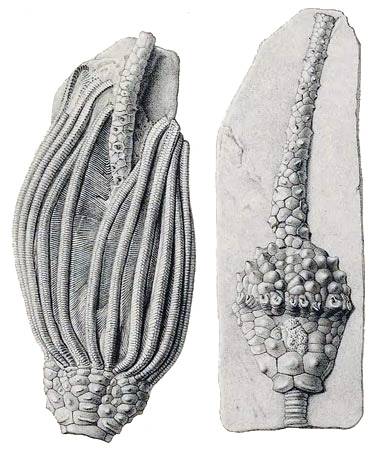
Obr 24 - Typický představitel kamerátního typu krinoida - Delocrinus Missouriensis Miller and Gurley, 1890 - Missouri, Mississippian (Visean). Všimněte si na ramenech vlevo hustých jemných pinnulí. Vpravo je zachovalý kalich s tegmenem a análním tubusem bez ramen. (printed for the Smithsonian Libraries in 1897).
K dalším významným vyhynulým zástupcům této podtřídy (i mimo Barrandien) patří především tyto rody: Apodasmocrinus, Calycanthocrinus, Cincinnaticrinus (Heterocrinus), Columbicrinus, Ectenocrinus, Halysiocrinus, Homocrinus, Isotomocrinus, Myelodactylus (Herpetocrinus), Ohiocrinus, Paracremacrinus, Ristnacrinus, Synbathocrinus, Triacrinus, Tryssocrinus, Zophocrinus.

Obr. 25 - ukázka stavby těla disparidní lilijice dle R. C. Mooreho

Obr. 26 - Typický představitel kamerátního typu krinoida - Synbathocrinus swallovi Hall, 1858.
Významné jsou především tyto rody: Caleidocrinus Jahn, 1893, Calceocrinus, Cremacrinus, Crinobrachiatus, Senariocrinus, Eucatillocrinus, Huxleyocrinus Donovan, 1985, Iocrinus Hall, 1866, Pisocrinus Rozhnov, 1981, Pygmaeocrinus Bouška 1947, Holycrinus, Pernerocrinus Bouška, 1947, Pachylocrinus by Lane et al. (2001), Encrinus Andreae, 1764.
Podtřída Cladida Moore & Laudon, 1943 †
(ordovik - spodní karbon)
Bohatě rozvětvená struktura paží, paže volně pohyblivé nad pohárem, pinulace ukončuje větvení ramen, dicyklický pohár, anální vak většinou dobře vyvinut. Největší rozšíření je v devonu a spodním karbonu. Známe asi 270 rodů. Rozeznáváme zde dva základní typy - primitivní kladidy (absence pinnulí) a pokročilé kladidy (s pinnulemi).

Obr. 27 - ukázka stavby těla kladidní lilijice dle R. C. Mooreho

Obr. 28 - Ukázka kladidních (Cladida) krinoidů. Vlevo pokročilá forma s velkými, hrubými pinnulemi, vpravo primitivní forma s větvenými rameny bez pinnulí. Mississippian (Visean).
Významné jsou v Barrandienu především tyto rody: Crotalocrinites Austin, 1842, Cupressocrinites Goldfuss, 1831 (1826), Cyathocrinites Miller, 1821, Gissocrinus Angelin, 1878, Jahnocrinus Jeakel, 1918, Pandoracrinus Jaekel, 1918 - sporný taxon ze šáreckého souvrství, Pernerocrinus Bouška, 1947, Polycrinus Jaekel, 1918, Syndetocrinus Kirk, 1933, Vosekocrinus Jaekel, 1918.
Podtřída Flexibilia von Zittel, 1895 †
(střední ordovik - svrchní perm)

Obr. 29 - ukázka stavby těla flexibilní lilijice dle R. C. Mooreho
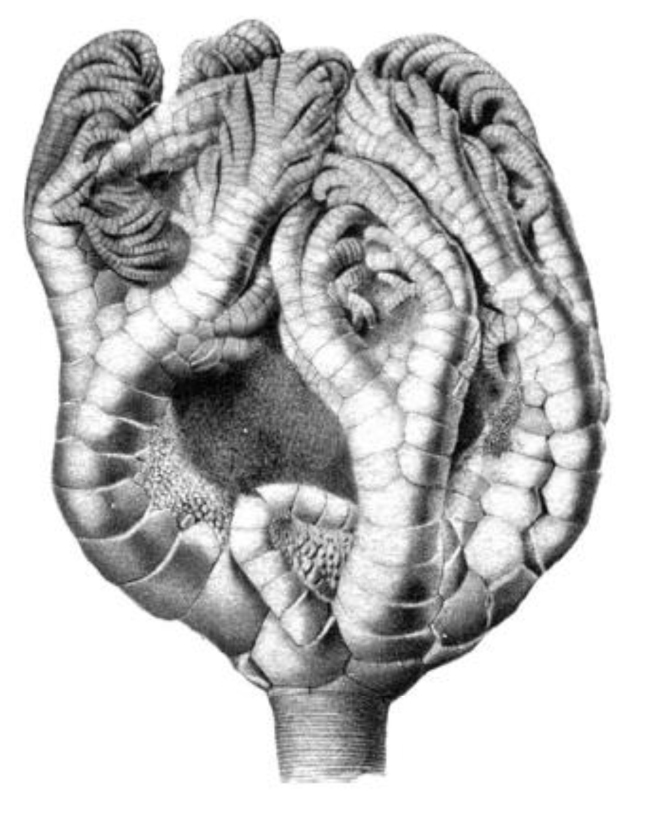
Obr. 30 - Ukázka charakteristického flexibilního krinoida čeledi Taxocrinidae Angelin 1878. Typické je pro tuto skupinu bohaté členění ramen.
Zdroje/použitá literatura:
- https://cs.wikipedia.org/wiki/Lilijice
- https://www.biomach.cz/biologie-zivocichua/ostnokozci
- https://zkamenely-svet.webnode.cz/
- https://muzeum.geology.cz/
- https://www.helsinki.fi/~mhaaramo/metazoa/deuterostoma/echinodermata/crinoidea/disparida.html
- Rudolf Prokop. Zkamenělý svět, vydala Práce v Praze, ISBN 80-208-0888-4, 1989
- https://www.asoldasthehills.org/Echinoderms.html
- Rudolf J. Prokop. Výzkum ostnokožců koněpruských vápenců Barrandienu (spodní devon, prag) - II, 2002
- Rudolf J. Prokop, Radana Slámová. NEW SPECIES OF CRINOIDS BASED ON THEIR COLUMNALS AND STEM FRAGMENTS (COL.), FROM THE LOWER DEVONIAN ZLÍCHOV LIMESTONE (BARRANDIAN AREA, THE CZECH REPUBLIC), 2012
- Dipl.-Ing. Joachim Hauser. Trybliocrinus collensis n.sp. - Ein neuer Vertreter von Trybliocrinus aus dem Emsium der La Vid Gruppe (Unterdevon) des Kantabrischen Gebirges (Nordspanien), 2010
- Rudolf J. Prokop. Zhistorie výzkumu ostnokožců českého staršího paleozoika, 2002
- Joachim Barrande. Système silurien du centre de la Bohème. Classe des Echinodermes. Famille des Crinoïdes, 1899
- Raymond C. Moore. Revision of Calceocrinidae, University of Kansas Publications, July 9, 1962
- FREDERICK H. C. HOTCHKISS, RUDOLF J. PROKOP, VÁCLAV PETR, Isolated vertebrae of brittlestars of the Family Klasmuridae Spencer, 1925 (Echinodermata: Ophiuroidea) in the Devonian of Bohemia (Czech Republic), Journal of the Czech Geological Society 44/3?4 (1999)
- A NEW DEVONIAN CRINOID FROM AUSTRALIA by DENIS E. B. BATES, 1972
- HANS HESS, WILLIAM I. AUSICH, CARLTON E. BRETT, MICHAEL J. SIMMS, Fossil Crinoids, Cambridge University Press 1999
- Zpravodaj České geologické společnosti - leden 2014 |18, Vydala © Česká geologická společnost, Praha 2014 - https://www.geologickaspolecnost.cz/soubory/zpravodaj/zpravodaj-cgs18.pdf
- von Dipl.-Ing. Joachim Hauser. Bactrocrinites hohenfelsensis HAUSER, 2006 - Ein neuer Vertreter des Taxon Bactrocrinites aus dem Junkerbergium (Mitteldevon) der Gerolsteiner Mulde (Rheinisches Schiefergebirge, Eifel), Bonn, mit 13 Textfiguren, 1 Tafel, 1 Tabelle und 18 Seiten Vorpubliziert im Internet am 24. November 2006
- RAYMOND C. MOORE and RUSSELL M. JEFFORDS, ECHINODERMATA - ARTICLE 9, CLASSIFICATION AND NOMENCLATURE OF FOSSIL CRINOIDS BASED ON STUDIES OF DISSOCIATED PARTS OF THEIR COLUMNS, The University of Kansas Paleontological Institute, JANUARY 26, 1968
- Hans Hess, Part T, Revised, Volume 1, Chapter 19: Paleoecology of Pelagic Crinoids, The University of Kansas, Paleontological Institute, 2010